")
")
Glossaire
Glossaires
| Terme | Définition |
|---|---|
| Durcissement | ♦ Stratégie dans laquelle les gestionnaires interviennent pour améliorer la résilience des ressources des aires protégées afin d’orienter les impacts des visiteurs. Le durcissement peut être physique, par la création d’une surface dure pour absorber les impacts physiques directs des activités des visiteurs (par ex. le revêtement d’un sentier fréquenté), ou métaphorique, auquel cas les gestionnaires durcissent l’expérience des visiteurs en les informant de leur impact négatif sur les conditions de ressources du fait de leur utilisation, afin qu’ils soient incités à réduire leurs impacts. |
| Dureté de l’eau | ♦ Teneur d'une eau en calcium. |
| Dynamique | ♦ Évolution spatio-temporelle des milieux naturels et semi-naturels, caractérisée par des changements dans la composition spécifique, la structure et le fonctionnement des écosystèmes. Pour les milieux terrestres, la végétation est le descripteur principal. |
| Dynamique de la végétation | ♦ Succession graduelle de formations ou de groupements végétaux qui traduit, au cours du temps, les changements physionomiques et floristiques de la végétation en un lieu donné. |
| Dynamique de restauration | ♦ Trajectoire conduisant à rétablir des propriétés et des caractéristiques naturelles (structure, fonction, composition spécifique) au sein d'un écosystème ou d'un habitat initialement dégradé. |
| Dynamique des biocénoses | ♦ Évolution temporelle de la structure d'une communauté en fonction des facteurs naturels ou de causes exogènes ayant induit une perturbation dans le système. |
| Dynamique des populations | ♦ Décrit les variations d'abondance dans l'espace et dans le temps des diverses espèces et en recherche les causes. Elle tente de comprendre les facteurs responsables des variations d'effectifs au sein d'une population. Ce terme diffère de celui de cinétique qui est l'étude descriptive des effectifs, de la densité et de la structure des populations et leurs transformations au cours du temps.
> La croissance d'une population est caractérisée par une série d'équations : N(t + Δt) = N(t) + B + I - E - D - H où N(t) : Nombre d'individus de l'année t
> Les éléments intervenant sur ces différents compartiments sont :
Les facteurs endogènes interviennent également mais, contrairement aux précédents, ils ne peuvent faire l'objet d'interventions. On y classe le taux de fécondité, la longévité, les comportements sexuels... En termes d'applications pratiques, comprendre la dynamique d'une population animale permet de gérer au mieux son exploitation éventuelle, ceci afin de ne prélever que le surplus et de laisser la population de reproducteurs à un niveau stable. > L'importance de la dynamique des populations, élément de base de la compréhension de la croissance de la population humaine et de manière générale, des tendances des espèces, a conduit à l'élaboration de modèles de plus en plus complexes. dN / dt = nN - mN = (n-m) N = rN où r : Taux d'accroissement intrinsèque (dit paramètre malthuséen),
Ce modèle aboutirait à une surpopulation d'une espèce dans la mesure où le taux d'accroissement est indépendant de l'effectif, ce qui est crédible sur le plan mathématique mais ne l'est pas sur le plan biologique. 2. Le modèle logistique dN / dt = rN (1 - N/K) où K : Capacité limite du milieu ; Le paramètre K est généralement interprété comme le nombre d'organismes pouvant être nourris ou abrités par les ressources. Il a une signification biologique pour les populations avec une forte interaction entre les individus contrôlant leur reproduction. No < K : population augmente et atteint un plateau. Il s'agit de la courbe logistique Lors d'une croissance logistique, la taille de la population tend vers la capacité de charge K. Tant que la taille de la population de dépasse pas K, la population continue de croître. Quand N excède K, la population décroît. K est la situation d'équilibre d'une population sous l'effet d'une croissance logistique. Ce modèle ne peut être utilisé qu'en fonction de certaines conditions :
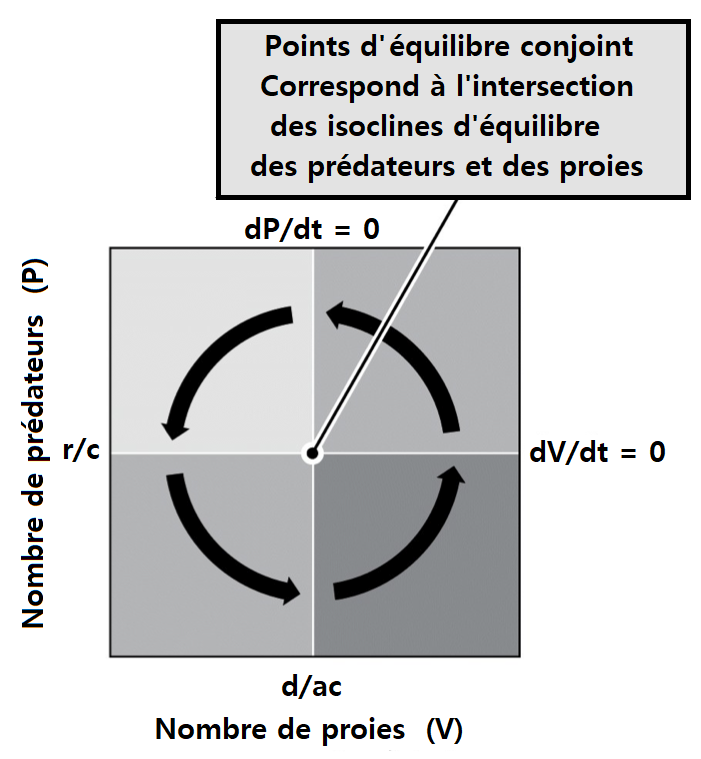
3. Modèle de Lotka-Volterra ou modèle proie-prédateur
dx / dt = rx - axy où a : Efficacité de prédation
> Dans les équations du modèle de Lotka-Volterra, la vitesse maximale d’accroissement d’une population de prédateurs se situe au moment de la densité maximale de ses proies reflétant le fait que le taux de multiplication du prédateur dépend essentiellement de la disponibilité de sa ressource almentaire. Réciproquement, la vitesse maximale de décroissance d’une population de proies se situe au moment de la densité maximale de prédateurs reflétant le fait que la cause de mortalité est la prédation. Les oscillations des deux populations sont donc de même fréquence mais décalées d’un quart de période.
♦ Équivalent étranger : Population dynamics. |
| Dynamique naturelle | ♦ Évolution dans le temps de la structure, du fonctionnement et de la composition spécifique des communautés végétales d'un stade initial, pionnier, vers un stade climacique. La dynamique naturelle est marquée par des perturbations et des processus naturels de grande envergure. |
| Dynamique progressive | ♦ Processus conduisant à une densification de la structure de la végétation et à la fermeture des milieux. |
| Dynamique régressive | ♦ Processus conduisant à une ouverture des milieux sous l'effet d'une perturbation naturelle ou anthropique. |
| Dynamique végétale | ♦ Succession dans le temps des espèces et groupements végétaux en un lieu donné. La dynamique progressive conduit des stades pionniers vers des stades plus natures dits climaciques, en général de types forestiers, en passant par tous les stades intermédiaires (prairie, formation arbustive). La dynamique est dite régressive lors du retour en arrière, dû à des causes naturelles (feu, glissement de terrain, érosion...) ou artificielles (défrichement, pâturage...). |
| Dysclimax | ♦ Communauté qui atteint un état d'équilibre qui ne sera jamais celui qui aurait dû être atteint dans des conditions normales, sous l'effet d'un facteur perturbateur naturel ou anthropique dont l'action est récurrente. Par exemple, dans les écosystèmes forestiers méditerranéens, les boisements de pins d'Alep sont des dysclimax dus à l'action récurrente du feu alors que, dans les conditions naturelles, le climax est représenté par des forêts de chênes verts ou de chênes pubescents selon les conditions locales. |
| Dysphotique | ♦ Se dit d'une zone profonde des écosystèmes aquatiques dans laquelle la lumière ne pénètre pas. En général, celle-ci est située en milieu océanique au-delà de profondeurs d'une centaine de mètres mais sa limite varie beaucoup selon la transparence du biotope aquatique considéré, allant de quelques dizaines de mètres à près de 300 mètres dans les cas extrêmes qui correspondent à des lacs hyper-oligotrophes. |
| Dystrophe -ique | ♦ 1. Qualifie un milieu en crise (rupture) de fonctionnement (blocage de la minéralisation due à un excès de matière organique). Une eau dystrophique est un plan d'eau peu profond qui contient beaucoup de matière organique et où l'eau fortement acide empêche la survie des poissons. |
| Dystrophie | ♦ État d'un milieu caractérisé par un apport excessif d'éléments minéraux nutritifs en raison de pollutions provoquant une eutrophisation accélérée des eaux. |