|
Bioévaluation environnementale
|
|
|
Biofilm
|
|
|
Biogène
|
♦ Qui est produit par les êtres vivants ou à l'opposé qui est nécessaire à la vie. On parlera, par exemple, d'une substance biogène ou des éléments biogènes pour désigner les éléments indispensables à la constitution de la matière vivante.
Synonyme : Biogénique.
♦ Équivalent étranger : Biogenic.
|
|
Biogénèse
|
♦ Ensemble des processus cosmologiques et physicochimiques à l'origine de la vie.
♦ Équivalent étranger : Biogenesis.
|
|
Biogéochimie
|
♦ Discipline scientifique qui traite de la transformation et du devenir de la matière, notamment de la matière organique et des éléments majeurs (carbone, azote, phosphore, silicium, etc.) dans la biosphère, par l'effet des processus biologiques, chimiques et géologiques.
♦ Équivalent étranger : Biogeochimistry.
|
|
Biogéochimique
|
♦ Un cycle biogéochimique est un processus qui caractérise la transformation de la matière et la circulation des éléments dans les écosystèmes et entre les divers compartiments de la biosphère. Les êtres vivants interviennent de façon déterminante dans ces cycles dont ils en sont les agents moteurs dans une phase ou une autre.
♦ Équivalent étranger : Biogeochemical.
|
|
Biogéocoenose
|
|
|
Biogéographie
|
♦ Étude scientifique de la distribution géographique des organes vivants. Elle définit la répartition spatio-temporelle des êtres vivants ainsi que les éléments et les causes qui conditionnent celle-ci. Elle s'intéresse aux causes actuelles de répartition des espèces (raisons climatiques par exemple) et aux causes anciennes d'ordre paléogéographique. La relation espèces/aire représente un des tous premiers modèles quantitatifs en biogéographie. Elle se présente sous la forme :
S' = A'z
où
• z décrit le rythme de rencontres des espèces sur une aire
• S' est la proportion des espèces espérées sur le site
• A' est la proportion de surface occupée par chaque type d'habitats
En utilisant cette équation, il est possible de prédire le nombre d'espèces observées si un pourcentage donné d'un type d'habitat est échantillonné, fourni par la valeur de z pour la végétation.
L'équation peut être utilisée pour formuler des objectifs de conservation pour des types d'habitats, pour déterminer la proportion de l'aire qui est requise pour représenter un pourcentage donné d'espèces :
Log A' = Log S'/z
♦ Équivalent étranger : Biogeography.
|
|
Biogéographie insulaire
|
♦ Branche de la biogéographie classique, elle vise, entre autres, à expliquer les facteurs conditionnant la richesse spécifique des communautés naturelles sur les îles. Elle trouve son origine dans les travaux du début des années 1960 et la publication de leurs résultats par McArthur et Wilson (1963 ; 1967), qui la développèrent en cherchant à prédire le nombre d'espèces qui pourraient s'implanter sur une île nouvellement créée. Cette théorie se fonde sur le postulat que le nombre d'espèces, dans une île non perturbée, est conditionné par l'immigration et l'extinction. L'immigration est dépendante de la distance de l'île au continent, ou à d'autres îles, servant de source de diffusion des espèces et des individus colonisateurs. Plus une île est isolée (par rapport à d'autres îles ou un continent), plus la probabilité qu'elle reçoive des espèces est faible.
La théorie reconnaît deux principes :
• La Théorie du non-équilibre prédit que le nombre d'espèces devrait augmenter au cours du temps ;
• La Théorie de l'équilibre prédit que le nombre d'espèces ne va pas changer au cours du temps mais que la composition de ces espèces va changer.
À tout moment, le nombre d'espèces sur une île est le résultat de l'équilibre entre les deux processus d'extinction et de colonisation. Le temps n'est donc pas un élément important car le nombre d'espèces présentes est lié plus à l'équilibre entre colonisation et extinction qu'à la durée pendant laquelle ces deux processus se développent. Si deux îles sont situées à une distance à peu près équivalente d'un continent, le taux de colonisation sera sensiblement identique
Les taux d'extinction, par contre, seront plus élevés sur l'île la plus petite. Aussi le nombre d'espèces à l'équilibre sera plus élevé sur la plus grande île. Le taux d'extinction d'une espèce cherchant à coloniser une île serait donc conditionné par la surface de l'île. La surface garantit qu'un habitat ne sera pas complètement détruit en cas d'événements importants, à moins d'une catastrophe majeure. Un plus grand nombre d'habitats augmente également le nombre d'espèces qui parviendront à s'implanter après l'immigration. Au cours du temps, les forces contraires de l'extinction et de l'immigration aboutissent à un niveau d'équilibre de la richesse spécifique. d'autres facteurs ont également été répertoriés comme ayant un rôle dans cet équilibre, le temps d'isolement de l'île, le climat, la localisation par rapport aux courants océaniques, la composition initiale en flore et en faune, les espèces les premières arrivées sur le site, le hasard d'une arrivée improbable et les activités humaines.
La biogéographie insulaire permet également de décrire la spéciation allopatrique qui se produit quand un nouveau pool génétique se développe à partir de la sélection naturelle intervenant dans un pool génétique isolé. Elle est également utile pour aborder la spéciation sympatrique, c'est-à-dire le concept selon lequel différentes espèces proviennent d'un même ancêtre dans une même aire.
L'immigration est déterminée par trois facteurs, la distance par rapport à la source, la diversité des espèces au niveau de la source et la probabilité qu'une espèce donnée puisse se disperser. L'extinction est déterminée par trois facteurs : la surface de l'île, la diversité des espèces sur l'île et la probabilité d'extinction de chaque espèce. Dans la version la plus simple du modèle, toutes les espèces ont des probabilités d'immigration et d'extinction égales. Dans la réalité, l'immigration est inversement liée à la distance séparant la source de l'île.
Eléments expliquant la probabilité d’extinction en fonction du taux d’immigration
> La probabilité d'extinction est faible si :
• r est élevé
• K est élevé (espèces de petite taille, généralistes, en bas de la chaine trophique)
• Les effectifs sont stables
Lorsque le taux d'immigration est faible :
- Si l'espèce est bon colonisateur, avec une différentiation génétique rapide
Lorsque le taux d'immigration est élevé :
• Si l'espèce est très bon colonisateur, avec une différentiation génétique lente
> La probabilité d'extinction est élevée si :
• le taux d'immigration est faible et l'espèce est mauvais colonisateur
• le taux d'immigration est élevé et l'espèces est bon colonisateur mais avec une différenciation génétique lente
Le nombre d'espèces sur une île répond donc à l'équation dite d'Arrhénius :
S = C Az
où S est le nombre d'espèces
A est la surperficie
C est une constante caractérisant la richesse biotique d'une aire
z est la pente, caractérisant la difficulté à atteindre une île
Quand une nouvelle île se forme, les espèces commencent à la coloniser et plus elles sont nombreuses et plus le rythme de colonisation tend à décliner. Le taux d'extinction commence à augmenter avec une augmentation de la densité. À un point donné, les deux processus s'équilibrent et le nombre d'espèces (S) sur l'île tend à se stabiliser.
Cette théorie est remise en cause pour plusieurs raisons :
• Elle se base sur le tryptique immigration, extinction, évolution sans prendre en compte différents éléments physiques comme la surperficie, la géologie, le climat qui vont les influencer sur des échelles temporelles et spatiales ;
• La surperficie n'est, en effet, pas forcément la cause première car elle peut conditionner le nombre et la surface d'habitats différents qui peuvent être utilisés par la faune. De grandes îles peuvent ainsi contenir plus d'habitats (ou de biotopes) que de petites îles, ce qui va conditionner le nombre d'espèces.
• La théorie ne fait référence qu'au nombre d'espèces et non aux densités de populations ou aux compositions d'espèces et ne se réfère qu'aux faunes insulaires dans leur globalité. Le développement historique des îles n'est pas pris en compte.
• La capacité de dispersion et les possibilités d'établir des populations ne sont pas prises en compte, tout comme la compétition et les interactions interspécifiques ;
• Enfin, la probabilité d'une colonisation réussie peut être déterminée par la présence de compétiteurs, directement par interférence ou par l'exploitation des ressources, l'une et l'autre pouvant conduire à réduire les populations les moins compétitrices et donc à leur extinction à terme. Aussi, le taux d'extinction ne serait donc pas uniquement lié aux habitats mais à la présence ou à l'absence d'autres espèces.

♦ Équivalent étranger : Island biogeography.
|
|
Biogéographie insulaire appliquée aux aires protégées
|
♦ Durant ces dernières décennies, la mise en place des aires protégées a été justifiée par la théorie de l'équilibre en biogéographie insulaire. Pour de nombreux chercheurs et gestionnaires, les aires protégées constituent des îlots au sein d'un ensemble différent - cultivé ou urbanisé - analogues aux îles du domaine maritime qui sont considérées comme un habitat favorable environné d'un habitat non favorable.
Scientifiques et conservateurs ont tenté d'établir un parallèle entre des îles isolées en mer ou dans un océan et des aires protégées isolées dans des espaces dégradés ou détruits. Les auteurs d'origine britannique ont appelé cette théorie (single large or several small [SLOSS], une seule grande ou plusieurs petites). Plusieurs principes de conservation furent exposés sur la base du postulat qu'il était préférable de disposer d'une aire protégée de grande superficie plutôt que de plusieurs aires protégées de faible superficie car les risques d'extinction sont ainsi réduits, en application de la théorie de l'équilibre. En d'autres termes, les communautés riches en espèces abritent tous types d'espèces, y compris celles rencontrées dans des communautés pauvres en espèces. Toutefois, ceci n'est pas nécessairement le cas pour les espèces endémiques ou pour les taxa qui ont été relégués dans des refuges de petite taille. Aussi, était-il important de considérer si une aire protégée est définie pour accueillir autant d'espèces que possible ou pour protéger un faible nombre d'espèces vulnérables. La théorie de l'équilibre de la biogéographie insulaire suggère que les systèmes d'aires protégées devraient contenir des éléments les plus étendus possibles, et que la distance entre les aires protégées devrait être la plus petite possible. Les plus grandes aires protégées devraient contenir les plus grands nombres d'espèces et donc contribuer au mieux à la préservation de la biodiversité.
Les résultats initiaux de l'application de cette théorie aboutirent à la définition de quatre stratégies :
• Les aires protégées doivent être aussi grandes que possible à l'image des grandes îles qui sont réputées moins sensibles aux risques d'extinction ;
• Des biotes uniques devraient être inclus dans autant d'aires protégées que possible et ces aires devraient être situées à proximité les unes des autres. Si l'une d'entre elles subit une extinction locale d'une espèce donnée, une autre aire protégée située à proximité pourrait fournir la base d'une reconquête ;
• Les aires protégées doivent être aussi circulaires que possibles pour minimiser l'effet de lisière. Ceci est fondé sur l'effet péninsule qui montre une tendance à une biodiversité plus basse. En réduisant les effets de lisière, on diminue les potentialités d'invasion par des espèces exotiques et par des perturbations extérieures ;
• Les gestionnaires et les conservateurs devraient prioriser les biotes avec la plus grande quantité d'endémisme et de vulnérabilité à la perte d'habitats.
> Cependant, créer une aire protégée de grande dimension n'est pas toujours facile en raison des contraintes physiques et de l'occupation du sol par les activités humaines. Si elle doit être fractionnée, la mise en place de corridors permettant de garantir la proximité et la connectivité entre les différentes sous-unités est nécessaire, avec cependant le risque que ces corridors puissent également être utilisés par des pathogènes et des maladies, ce qui nécessite donc une définition très stricte de leur emplacement. Bien qu'il soit accepté que le nombre d'espèces d'une aire protégée est une fonction croissante de la surface, des prédictions plus spécifiques ont donné lieu à un large débat notamment parce que l'utilisation de la courbe existant entre la surface et le nombre d'espèces ne peut être complète en raison d'éléments divers biotiques et abiotiques qui viennent interférer. Enfin, la théorie ne parle pas de l'identité des espèces qui risquent de s'éteindre.
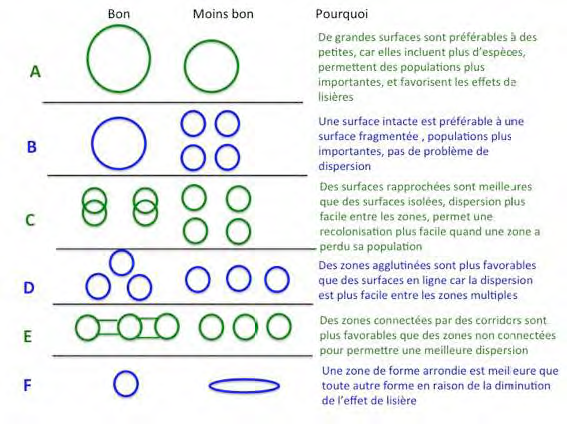 Représentation schématique de la théorie du SL Représentation schématique de la théorie du SL
Les grandes réserves contiennent plus d'espèces, perdent des espèces plus lentement et souffrent moins des effets délétères de l'isolement des habitats que les réserves plus petites. Les réserves isolées des autres aires d'habitats par des paysages très vastes mais inhospitaliers, vont accueillir moins d'espèces que celles qui sont plus proches de paysages plus naturels. La fragmentation des habitats affecte les espèces de différentes façons. Certaines déclinent fortement ou disparaissent dans les fragments tandis que d'autres demeurent stables et que d'autres enfin augmentent, parfois très fortement.
Les espèces les plus sensibles à l'extinction ne se rencontrent souvent que dans les grandes réserves, favorisant ainsi la stratégie d'une seule grande réserve, bien que de petites réserves, éparpillées dans une région, peuvent abriter des espèces à faible répartition qui, sans cela, ne seraient pas protégées. Les perturbations écologiques, comme le feu ou les invasions d'insectes peuvent également jouer un rôle. Pour cette raison, la superficie d'une aire protégée devrait être toujours plus grande que la plus grande perturbation afin de permettre la reconquête interne par les espèces survivantes.
Enfin, les aires protégées ne ressemblent pas tout à fait à des îles, ce qui réduit la possibilité d'utiliser la théorie de la biogéographie insulaire. Leurs limites sont souvent plus liées à des contraintes économiques et politiques qu'à l'intégration de l'ensemble d'un écosystème. Parfois, il n'y a pas de choix, le contexte politique imposant d'accepter n'importe quelle surface, pourvu qu'une aire protégée puisse être définie. Aussi, la réponse à la question « une grande aire protégée plutôt que plusieurs petites » dépend des objectifs, des espèces que l'on cherche à préserver et pourquoi, et enfin des ressources disponibles.
 Représentation des empires biogéographiques Représentation des empires biogéographiques
♦ Équivalent étranger : Island biogeography adapted to protected areas.
|
|
Biogéographique
|
|
|
Biogéomorphologie
|
|
|
Biogéosphère
|
♦ Zone superficielle de la planète où se trouvent tous les êtres vivants.
♦ Équivalent étranger : Biogeosphere.
|
|
Bioindicateur
|
♦ Peut se définir comme une espèce ou un groupe d'espèces végétales ou animales dont les caractéristiques observées (occurrence, abondance, biomasse, caractéristiques ou « traits » biologiques) fournissent une indication sur le niveau de dégradation du milieu.
Il fait donc l'objet de mesures permettant d'indiquer la présence ou les effets des polluants. Un bioindicateur est un outil d'évaluation de la qualité de l'environnement. Il peut être considéré comme complémentaire (notamment pour la répartition spatiale de la pollution, la caractérisation des synergies, la pollution graduelle...) des réseaux de mesures physico-chimiques pour les raisons suivantes :
- installation rapide et facile avec une faible infrastructure ;
- grande souplesse au niveau de la modification, extension ou déplacement de ces réseaux ;
- faible coût, tant au niveau de l'investissement que du suivi ;
- réalisation d'études préliminaires permettant ensuite de choisir avec précision les emplacements les mieux adaptés à l'implantation de capteurs physico-chimiques ;
- densification à faible coût des réseaux de capteurs physico-chimiques ;
- détection des polluants nouveaux ou accidentels non pris en compte par les capteurs en place.
> Les espèces bioindicatrices idéales sont :
- opportunistes, tolérantes aux perturbations de l'environnement ;
- sédentaires et abondantes sur les sites à étudier ;
- faciles à récolter et à identifier ;
- accumulent et ne régulent pas les contaminants ;
- reflètent les variations de la qualité chimique du milieu.
> Les bioindicateurs peuvent être utilisés pour :
- émettre des signaux précoces de problèmes environnementaux ;
- identifier les relations de cause à effet entre facteurs d'altération et effets biologiques ;
- évaluer l'état de stress global de l'environnement à travers différentes réponses d'organismes indicateurs ;
- évaluer l'efficacité de mesures réparatrices sur la santé des systèmes biologiques.
> Leur utilisation présente certaines limites :
- disponibilité non permanente pour certaines espèces végétales ;
- possibilités d'interférences par d'autres stress biotiques ou abiotiques dans les réponses ;
- fourniture d'informations essentiellement relatives aux niveaux de pollution atmosphérique (aspect qualitatif) ;
- pas encore de véritable reconnaissance ni de normalisation des méthodes aussi bien sur la mise en œuvre que sur l'interprétation des résultats (échelle de correspondance).
Une métrique de bioindication est une grandeur calculée qui décrit certains aspects de la structure, de la fonction ou de tout autre caractéristique des assemblages biologiques et qui change de valeur en réponse à une modification de l'impact des activités humaines.
♦ Équivalent étranger : Bioindicator.
|
|
Bioinformatique
|
♦ Discipline émergente de la recherche qui se place à l'interface de la biologie et de l'informatique, consistant en l'utilisation de l'outil informatique pour traiter les données de la biologie. Les moyens informatiques sont naturellement utilisés pour le stockage ou la gestion des données mais également pour l'interprétation de ces données. Par opposition aux classiques manipulations in vivo ou in vitro pratiquées en laboratoire, on parle d'expériences « in silico » (néologisme formé à partir de l'anglais silicon).
♦ Équivalent étranger : Bioinformatics.
|
")
")